 Medi TrendmmmmmMedi Trendmmmmm
Medi TrendmmmmmMedi TrendmmmmmBasen und Nukleotide
Die Nucleinsäuren spielen eine zentrale Rolle bei der Speicherung und Expression der genetischen Information. Man unterscheidet zwei Hauptklassen: Desoxyribonucleinsäure (DNA) dient ausschließlich der Informationsspeicherung, während Ribonucleinsäuren (RNAs) an den meisten Schritten der Genexpression und der Proteinbiosynthese beteiligt sind. Alle Nucleinsäuren sind aus Nucleotid-Bausteinen aufgebaut, die wiederum aus einer Base, einem Zucker und einem Phosphat-Rest bestehen. DNA und RNA unterscheiden sich in der Art des Zuckers und in einer der Basen.
Basen
Die Nucleinsäure-Basen sind aromatische Heterocyclen, die sich entweder vom Pyrimidin oder vom Purin ableiten. Fünf dieser Basen sind als Hauptkomponenten der Nucleinsäuren aller Lebewesen gemeinsam. Die Purin-Basen Adenin (Abkürzung Ade, nicht A) und Guanin (Gua) sowie die Pyrimidin-Base Cytosin (Cyt) kommen in RNA und DNA vor. Dagegen findet sich Uracil (Ura) nur in RNA. In der DNA nimmt Thymin (Thy), das 5-Methyl-Derivat des Uracils, dessen Platz ein. In der DNA höherer Organismen tritt in kleinen Mengen auch 5-Methylcytosin auf. Eine große Zahl weiterer abgewandelter Basen findet man in der tRNA und in anderen RNA-Arten.
Nucleoside, Nucleotide
Verknüpft man eine Nucleinsäure-Base mit Ribose oder 2-Desoxyribose, erhält man ein Nucleosid. So entsteht z.B. aus Adenin und Ribose das Nucleosid Adenosin (Abkürzung A). Die entpsrechenden Derivate der anderen Basen heißen Guanosin (G), Uridin (U), Thymidin (T) und Cytidin (C). Ist 2-Desoxyribose die Zuckerkomponente, entsteht ein Desoxynucleosid, z.B. 2'-Desoxyadenosin (dA). In der Zelle ist die 5' OH-Gruppe der Zuckerkomponente der Nucleoside i.a. mit Phosphorsäure verestert. Aus 2'-Desoxythymidin (dT) entsteht so 2'-Desoxythymidin-5'monophosphat (dTMO), ein DNA-Baustein. Wird der 5'-Phosphat-Rest über Säureanhydrid-Bindungen mit weiteren Phosphat-Resten verknüpft, erhält man Nucleosid-di- und triphosphate, z.B. ADP und ATP, wichtige Coenzyme des Energiesoffwechsels. Alle diese Nucleosid-Phosphate faßt man unter dem Begriff Nucleotide zusammen. In Nucleosiden undd Nucleotiden liegen die Pentose-Reste in der Furanose-Form vor. Zucker und Base sind durch eine N-glykosidische Bindung zwischen C-1' des Zuckers und N-9 des Purin-Rings bzw. N-1 des Pyrimidin-Rings verknüpft. Diese Bindung ist stets ß-konfiguriert.
Basenpaarung in der DNA
Erste Hinweise auf die Existenz solcher Wechselwirkungen brachte die Beobachtung, daß in jeder Art von DNA der Gehalt an Adenin und Thymin annährend gleich sind. Dasselbe gilt für Guanin und Cytosin. Dagegen ist das Verhältnis der Summe von Ade und Thy zur Summe von Gua und Cyt von Organismus zu Organismus verschieden. Das 1953 formulierte Modell der DNA-Struktur erklärt diese konstanten Basenverhältnisse: Intakte DNA besteht aus zwei Polydesoxynucleotid-Moleküken (Strängen). Jede Base im einen Strang ist mir einer komplementären Base im anderen Strang durch H-Brücken verknüpft. Dabei ist Adenin komplementär zu Thymin, und Guanin komplementär zu Cytosin. An jedem Basenpaar ist also eine Purin- und eine Pyrimidin-Base beteiligt. Die Komplementarität von A und T bzw. G und C wird verständlich, wenn man die möglichen H-Brücken zwischen den Basen betrachetet. Also Donoren kommen Amino-Gruppen (Ade, Cyt, Gua) und Ring-NH-Gruppen (Gua, Thy) in Betracht. Mögliche Akzeptoren sind Carbonylsauertoff-Atome (Thy, Cyt, Gua) und Stickstoff-Atome im Ring. In A-T Paaren können sich deshalb zwei, im G=C Paaren sogar drei lineare und damit besonders stabile Brücken ausbilden. Das in RNA statt Thymin vorkommende Uracil verhält sich bei der Basenpaarung genauso wie Thymin.
DNA-Struktur
Basenpaarungen können sich in DNA über Millionen von Bausteinen erstrecken. Sie sind allerdings nur möglich, wenn die Polarität der beiden Stränge unterschiedlich ist, d.h. wenn sie unterschiedliche Richtungen haben. Außerdem müssen die beiden Stränge in Form einer Doppelhelix umeinander gewunden sein. RNA kann wegen sterischer Hinderung durch die 2'-OH Gruppe der Ribose-Reste keine derartige Doppelhelix ausbilden. Ihre Struktur ist deshalb weniger regelmäßig als die von DNA. Da die beiden Stränge nur durch nichtkovalente Wechselwirkungen zusammengehalten werden, kann man die DNA-Doppelhelix durch Erhitzen oder durch Inkubation in alkalischer Lösung leicht in die Einzelstränge trennen (Denaturierung). Beim langsamen Abkühlen der weitgehend ungeordenetet Einzelstränge bildet sich durch Basenpaarung die ursprünglische Dopelhelix wieder zurück (Renaturierung). Die De- und Renaturierung von DNA spielt in der Gentechnik eine wichtige Rolle. Funktionell sind die beiden DNA-Stränge nicht äquivalent: Der codierende Strang (Matrizen-Strang) ist derjenige, der bei der Transkription abgelesen wird. Er ist komplementär zu RNA. Der nichtcodierende Strang hat abgesehen vom Austausch von T gegen U, die gleiche Sequenz wie die RNA. Man ist übereingekommen, Gegensequenzen als Sequenz des nichtcodierenden Stranges der DNA in 5' -> 3' - Richtgung anzugeben. Liest man die Codons in dieser Weise, erhält man mit Hilfe des genetischen Codes direkt die Proteinsequenz in der üblichen Leserichtung, also vom N-Terminus zum C-Terminus.
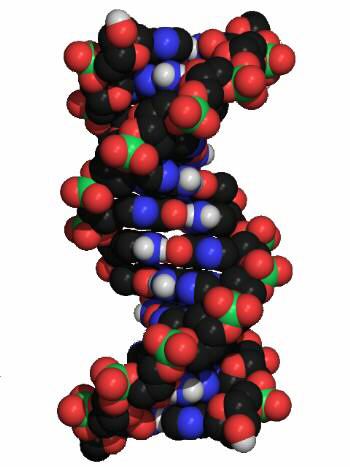
B-DNA
Durch Untersuchungen an synthetischen DNA-Molekülen fand man, daß DNA verschiedene Konformationen einnehmen kann. Die weitaus häufigste Form ist die B-DNA. Wie bereits erwähnt, besteht sie aus zwei gegenläufigen Polydesoxynucleotid-Strängen, die als rechtsgängige Doppelhelix umeinander gewunden sind. Das Rückgrat dieser Stränge wird von Desoxyribose- und Phosphat-Resten gebildet, die durch Phosphorsäurediester-Bindungen verknüpft sind. Die aromatischen Ringe der Basen sind in einem Abstand von 0,34 nm annähernd rechtwinklig zur Helixachse gestapelt. Jede Base ist gegenüber der vorhergehenden um einen Winkle von 35° gedreht. Eine volle Windung der Doppelhelix (360°) enthält daher etwa 10 Basenpaare, die Ganghöhe der Helix beträgt 3,4 nm. Zwischen den Rückgraten der beiden Einzelstränge gibt es zwei unterschiedliche Furchen. Die große Furche ist oben und unten sichtbar, die schmalere kleine Furche in der Mitte. DNA-bindenden Proteine treten meist mit den besser zugänglichen Basen im Bereich der großen Furche in Wechselwirkung. Unter bestimmten Bedingengen kann DNA in die A-Konformation übergehen. In dieser Anordnung bleibt die rechtsgängige Doppelhelix erhalten, die Basen sind aber nicht senkrecht zur Helix-Achse angeordent, wie in der B-Form, sondern geneigt. In der Z-Konformation, die in GC-reichen Teilsequenzen innerhalb von B-DNA auftreten kann, sind die Anordnungen der Nucleotide völlig verschieden: Die Doppelhelix ist hier linksgängig, außerdem hat das Rückgrat der Stränge eine charakteristische Zickzack-Form (daher Z-DNA).
RNA
Die Ribonucleinsäuren (RNAs) sind Polymere aus Nucleosid-phosphat-Bausteinen, die durch Phosphorsäuredieester-Bindungen verknüpft sind. Als Basen enthalten sie vor allem Uracil, Cytosin, Adenin und Thymin, aber auch viele ungewöhnliche und modfizierte Basen sind in RNAs zu finden. RNAs sind an allen Teilschritten der Genexpression und Proteinbiosynthese beteiligt. Im Gegensatz zur DNA können RNAs keine ausgedehnten Doppelhelices bilden. Bei ihnen erstrecken sich Basenpaarungen meist nur über wenige Reste. Deshalb entstehen häufig Substrukturen, die in zweidimensionaler Darstellung fingerförmig oder kleeblattartig erscheinen. In ihnen sind gepaarte Stamm-Bereiche (stem) durch Schleifen (loop) verbunden. Große RNAs wie die ribosomale16S-rRNA enthalten zahlreiche "stem-and-loop" Bereiche dieser Art. Diese Abschnitte sind wieder dreidiemsional gefaltet, d.h. RNAs haben wie Proteine eine Tertiärstruktur. Bisher kennt man Tertiärstrukturen nur von kleinen RNAs, vor allem von tRNA. Die RNAs der Zelle unterschieden sich erheblich in Größe, Struktur und Lebensdauer. Den bei weitem größten Anteil hat die ribosomale RNA (rRNA), die in mehreren Formen struktureller und funktioneller Besdanteil der RIbosomen ist. Ribosomale RNA wird im Nucleolus durch Transkription aus DNA erzeugt und dort aus prozessiert und mit Proteinen zu Ribosomen-Untereinheiten zusammengebaut. Messanger-RNAs (mRNA) übertragen die genetische Information vom Zellkern ins Cytoplasma. Auch ihre Transkripte werden noch im Zellkern stark verändert (mRNA-Reifung). Da mRNAs am Ribosom Codon für Codon abgelesen werden müssen, dürfen sie keine stabile Tertiärstruktur ausbilden. Dies wird auch dadurch erreicht, daß sich RNA-bindende Proteine anlagern, die Basenpaarungen verhindern. Wegen der unterschiedlichen Menge an Information, die sie tragen, sind auch die Längen der mRNAs sehr verschieden. Ihre Lebensdauer ist meist kurz, da sie nach der Translation rasch wieder abgebaut werden.
tRNA
Die Transfer-RNAs (tRNAs) fungieren in der Translation als Bindeglieder zwischen den Nucleinsäuren und den Proteinen. Es sind kleine RNA-Moleküle aus 70-90 nt, die mit ihrem Anticodon durch Basenpaarung bestimmte mRNA-Codons erkennen. Gleichzeitig tragen sie am 3'-Ende diejenige Aminosäure, die nach dem genetischen Code dem betreffenden mRNA-Codon zugeordnet ist. Zahlreiche Basenpaarungen, die zum Teil vom üblichen Schema abweichen, stabilisieren die Konformation des Moleküls.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}